RESEARCH LINES

1. Effects of anthropogenic perturbations on nutrient cycling.
The biogeochemical cycles of carbon (C), nitrogen (N) and phosphorus (P) are interlinked by primary production, respiration and decomposition in terrestrial ecosystems. These biogeochemical cycles are the foundation of multiple ecosystem services such as soil fertility, food and fibre production and climate regulation (e.g., C storage), which are critical to support life on Earth. Increasing evidence exists to substantiate the claim that predicted global environmental changes such as those from climate change and land use intensification will alter C, N and P content and stoichiometry in terrestrial ecosystems. Improving our understanding of the responses of ecological stoichiometry to global change and other human legacies is of paramount importance to determine ecosystem responses and develop adaptation strategies for climate change and land use intensification.
Example 1. Our work suggest that any predicted increase in aridity, a common expression of climate change in drylands worldwide, will reduce the concentration of N and C in global drylands, but increase that of P. These changes would uncouple the C, N and P cycles in drylands and could negatively affect the provision of key ecosystem services provided by these ecosystems. To read more about this work, please have a look on our manuscript published in Nature (Delgado-Baquerizo et al. 2013) about how increases in aridity will decouple nutrient cycling in global drylands.
Example 2. More recently, we found that human legacies such as those derived from plant domestication have largely altered crop C:N:P stoichiometries; being the latitude of the domestication sites a major driver of these shifts in stoichiometry. Our findings draw attention to evolutionary effects of domestication legacies on plant and soil stoichiometry and related ecosystem services (e.g. current plant yield and soil fertility). To read more about this work, please have a look on our manuscript published in Ecology Letters (Delgado-Baquerizo et al. 2016) about how domestication of plants have shifted nutrient stoichiometry across multiple crop species.
2. Link between microbial communities and ecosystem functions
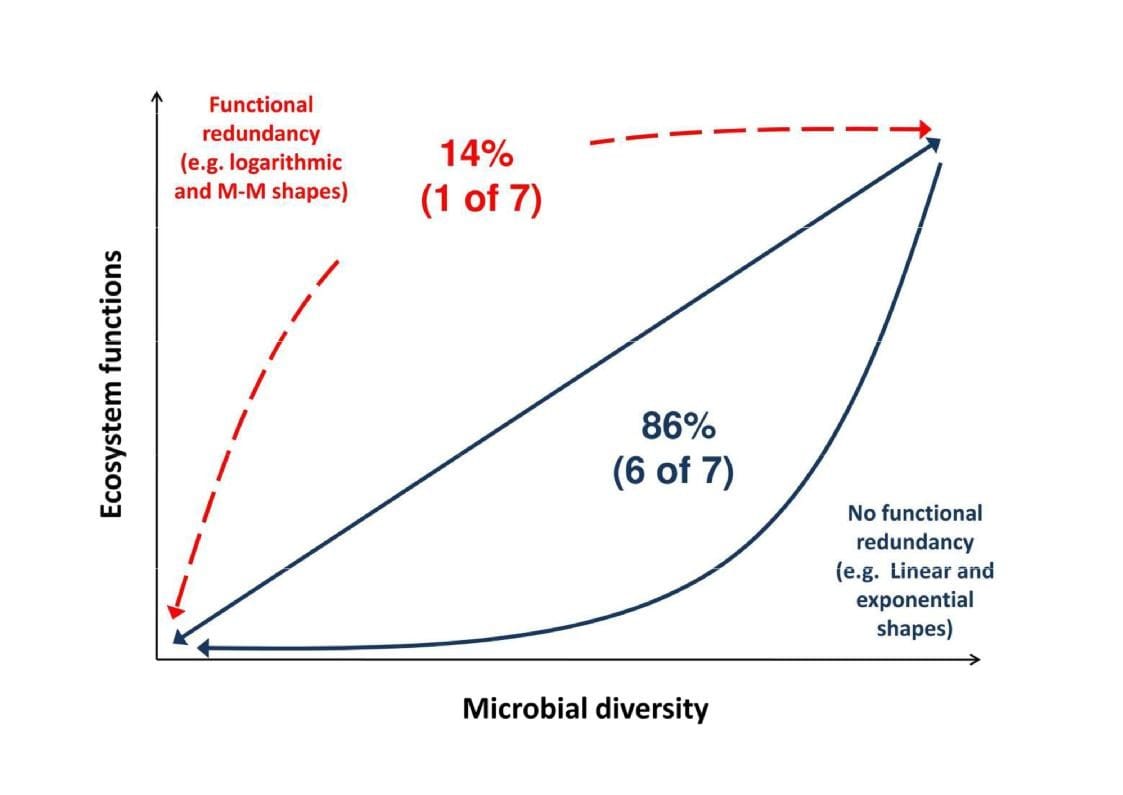
Unlike plants, we have limited knowledge of the relationship between microbial diversity and ecosystem functioning (aka microbial BEF), particularly in terrestrial environments. Microbial communities play key roles in maintaining multiple ecosystem functions and services simultaneously (‘multifunctionality’ hereafter), including nutrient cycling, primary production, litter decomposition and climate regulation. A growing number of studies suggest that microbial diversity enhances ecosystem functioning. However, unless there is a substantial functional redundancy in microbial communities, any loss in microbial diversity would likely alter the capacity of microbes to support ecosystem functions. Global environmental drivers such as climate change (e.g. see our paper Maestre et al. 2015) and land use intensification (e.g. see our paper Delgado-Baquerizo et al. 2016) are impacting microbial diversity. In order to evaluate the global consequences of shifting microbial diversity on ecosystem functioning, it is critical that we determine the shape of the microbial BEF relationship and to assess whether this relationship is maintained at the large scale in natural ecosystems from the “real world”.
Example 1. Our recent work suggest that unlike the positive but decelerating BEF relationship observed most frequently in plants and animals, ecosystem functions relate to bacterial diversity in a non-redundant fashion (e.g. exponentially and/or linearly). Reduced bacterial diversity, even after accounting for biomass, caused a decrease in broad (i.e., CO2 respiratory fluxes: conducted by many microbial communities) and specialized (toxin degradation: conducted by few microbial communities) functions in all evaluated cases. To read more about this work, please have a look on our manuscript published in Journal of Ecology (Delgado-Baquerizo et al. 2016) where we assess the shape of the bacterial BEF relationship in multiple freshwater ecosystems from Australia and United Kingdom.
Example 2. Recently, we report that soil microbial diversity positively relates to multifunctionality in terrestrial ecosystems across the globe. Our findings provide empirical evidence that any loss in microbial diversity will likely reduce multifunctionality, negatively impacting the provision of services such as climate regulation, soil fertility and food and fibre production by terrestrial ecosystems. To read more about this work, please have a look on our manuscript published in Nature Communications (Delgado-Baquerizo et al. 2016) where we evaluate the relationship between microbial diversity and ecosystem multifunctionality in terrestrial ecosystem at the global scale.
Unlike plants, we have limited knowledge of the relationship between microbial diversity and ecosystem functioning (aka microbial BEF), particularly in terrestrial environments. Microbial communities play key roles in maintaining multiple ecosystem functions and services simultaneously (‘multifunctionality’ hereafter), including nutrient cycling, primary production, litter decomposition and climate regulation. A growing number of studies suggest that microbial diversity enhances ecosystem functioning. However, unless there is a substantial functional redundancy in microbial communities, any loss in microbial diversity would likely alter the capacity of microbes to support ecosystem functions. Global environmental drivers such as climate change (e.g. see our paper Maestre et al. 2015) and land use intensification (e.g. see our paper Delgado-Baquerizo et al. 2016) are impacting microbial diversity. In order to evaluate the global consequences of shifting microbial diversity on ecosystem functioning, it is critical that we determine the shape of the microbial BEF relationship and to assess whether this relationship is maintained at the large scale in natural ecosystems from the “real world”.
Example 1. Our recent work suggest that unlike the positive but decelerating BEF relationship observed most frequently in plants and animals, ecosystem functions relate to bacterial diversity in a non-redundant fashion (e.g. exponentially and/or linearly). Reduced bacterial diversity, even after accounting for biomass, caused a decrease in broad (i.e., CO2 respiratory fluxes: conducted by many microbial communities) and specialized (toxin degradation: conducted by few microbial communities) functions in all evaluated cases. To read more about this work, please have a look on our manuscript published in Journal of Ecology (Delgado-Baquerizo et al. 2016) where we assess the shape of the bacterial BEF relationship in multiple freshwater ecosystems from Australia and United Kingdom.
Example 2. Recently, we report that soil microbial diversity positively relates to multifunctionality in terrestrial ecosystems across the globe. Our findings provide empirical evidence that any loss in microbial diversity will likely reduce multifunctionality, negatively impacting the provision of services such as climate regulation, soil fertility and food and fibre production by terrestrial ecosystems. To read more about this work, please have a look on our manuscript published in Nature Communications (Delgado-Baquerizo et al. 2016) where we evaluate the relationship between microbial diversity and ecosystem multifunctionality in terrestrial ecosystem at the global scale.

3. Patterns and mechanisms driving soil microbial communities
Given the functional role of soil microbes, understanding soil microbial diversity patterns is of paramount importance to establish effective policies to preserve microbial diversity hotspots and the key ecosystem functions and services that soil microbes provide. However, there are still important gaps in our understanding of the patterns and drivers of soil microbial diversity in terrestrial ecosystems. Some of these drivers include, but are limited to, biotic and abiotic factors such as spatial influence, climate, salinity, soil pH and organic matter. More studies are still needed, however, to provide an integrative view of how these biotic and abiotic factors influence soil bacterial diversity from the local to the global scales.
Example 1. In our recent meta-analysis in Ecological Monograph (Delgado-Baquerizo et al. 2016), we provided a novel integrative view of how climate and soil factors influence soil bacterial diversity at the global scale, which is critical to improve ecosystem and earth system simulation models and for formulating sustainable ecosystem management and conservation policies.
Example 2. The activity of mammalian engineers on soil showed to modify soil microbial communities and ecosystem functions at the local scale. Native species from Australia such as Echidna had positive effects on soil processes (e.g. decomposition) and showed a positive effect on the relative abundance of particular microbial taxa such as Proteobacteria at the expense of other taxa such as Acidobacteria. To read more about this work, please take a look to Eldridge et al. (2016).
Given the functional role of soil microbes, understanding soil microbial diversity patterns is of paramount importance to establish effective policies to preserve microbial diversity hotspots and the key ecosystem functions and services that soil microbes provide. However, there are still important gaps in our understanding of the patterns and drivers of soil microbial diversity in terrestrial ecosystems. Some of these drivers include, but are limited to, biotic and abiotic factors such as spatial influence, climate, salinity, soil pH and organic matter. More studies are still needed, however, to provide an integrative view of how these biotic and abiotic factors influence soil bacterial diversity from the local to the global scales.
Example 1. In our recent meta-analysis in Ecological Monograph (Delgado-Baquerizo et al. 2016), we provided a novel integrative view of how climate and soil factors influence soil bacterial diversity at the global scale, which is critical to improve ecosystem and earth system simulation models and for formulating sustainable ecosystem management and conservation policies.
Example 2. The activity of mammalian engineers on soil showed to modify soil microbial communities and ecosystem functions at the local scale. Native species from Australia such as Echidna had positive effects on soil processes (e.g. decomposition) and showed a positive effect on the relative abundance of particular microbial taxa such as Proteobacteria at the expense of other taxa such as Acidobacteria. To read more about this work, please take a look to Eldridge et al. (2016).

4. Biocrusts: drivers of ecosystem functions and major modulators of ecosystem responses to global change
Soil communities dominated by mosses, lichens and cyanobacteria (biocrusts hereafter) are common biotic components of boreal, arctic, temperate and dryland ecosystems worldwide. These communities support a wide range of ecosystem functions, including soil stability, carbon (C) and nitrogen (N) fixation, carbon dioxide flux, and N mineralization (see Castillo-Monroy et al. 2010 and Delgado-Baquerizo et al. 2015 for examples). Given the global distribution of biocrusts and their key functional roles in the ecosystems where they are prevalent, understanding how climate change will affect the capacity of these organisms to maintain multiple ecosystem functions simultaneously is critical for formulating sustainable natural resource management and conservation policies in drylands worldwide.
Example 1. Using samples from three continents (North America, Europe and Australia), we evaluated how the increase in aridity predicted by climate change will alter the capacity of biocrust-forming mosses to modulate multiple ecosystem processes related to C, N and P cycles. Our findings provide strong evidence that the maintenance of biocrusts is crucial to buffer negative effects of climate change on multifunctionality in global drylands. To read more about this work, please take a look to our manuscript in New Phytologist: Delgado-Baquerizo et al. (2016).
Example 2. Our recent work in Soil Biology and Biogeochemistry suggests that identity of biocrust species and microbial communities drives the response of soil multifunctionality to simulated global change. These findings highlight the importance of preserving biocrusts as hotspots of microbial genetic resources and ecosystem functioning in drylands. To read more about this work, please have a look to Liu et al. (2017).
Soil communities dominated by mosses, lichens and cyanobacteria (biocrusts hereafter) are common biotic components of boreal, arctic, temperate and dryland ecosystems worldwide. These communities support a wide range of ecosystem functions, including soil stability, carbon (C) and nitrogen (N) fixation, carbon dioxide flux, and N mineralization (see Castillo-Monroy et al. 2010 and Delgado-Baquerizo et al. 2015 for examples). Given the global distribution of biocrusts and their key functional roles in the ecosystems where they are prevalent, understanding how climate change will affect the capacity of these organisms to maintain multiple ecosystem functions simultaneously is critical for formulating sustainable natural resource management and conservation policies in drylands worldwide.
Example 1. Using samples from three continents (North America, Europe and Australia), we evaluated how the increase in aridity predicted by climate change will alter the capacity of biocrust-forming mosses to modulate multiple ecosystem processes related to C, N and P cycles. Our findings provide strong evidence that the maintenance of biocrusts is crucial to buffer negative effects of climate change on multifunctionality in global drylands. To read more about this work, please take a look to our manuscript in New Phytologist: Delgado-Baquerizo et al. (2016).
Example 2. Our recent work in Soil Biology and Biogeochemistry suggests that identity of biocrust species and microbial communities drives the response of soil multifunctionality to simulated global change. These findings highlight the importance of preserving biocrusts as hotspots of microbial genetic resources and ecosystem functioning in drylands. To read more about this work, please have a look to Liu et al. (2017).
